Митохондрии есть у всех типов эукариотных клеток (рис. 1). Они имеют вид либо округлых телец, либо палочек, реже - нитей. Их размеры колеблются от 1 до 7 мкм. Число митохондрий в клетке составляет от нескольких сотен до десятков тысяч (у крупных простейших).
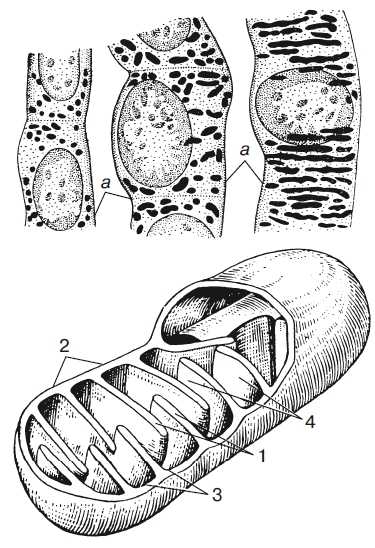
Рис. 1. Митохондрии. Вверху — митохондрии (?) в мочевых канальцах, видимые в световом микроскопе. Внизу — трехмерная модель организации митохондрии: 1 — кристы; 2 — внешняя мембрана; 3 — внутренняя мембрана; 4 — матрикс
Митохондрия образована двумя мембранами — внешней и внутренней, между которыми расположено межмембранное пространство. Внутренняя мембрана образует множество впячиваний — крист, представляющих собой либо пластины, либо трубочки. Такая ее организация обеспечивает огромную площадь внутренней мембраны. На ней располагаются ферменты, обеспечивающие преобразование энергии, заключенной в органических веществах (углеводах, липидах), в энергию АТФ, необходимую для жизнедеятельности клетки. Следовательно, функция митохондрий — участие в энергетических клеточных процессах. Именно поэтому большое количество митохондрий присуще, например, мышечным клеткам, выполняющим большую работу.
Пластиды. В растительных клетках обнаруживаются особые органоиды — пластиды, имеющие чаще веретеновидную или округлую форму, иногда более сложную. Различают три вида пластид — хлоропласты (рис. 2), хромопласты и лейкопласты.
Хлоропласты отличаются зеленым цветом, который обусловлен пигментом — хлорофиллом, обеспечивающим процесс фотосинтеза, т. е. синтеза органических веществ из воды (Н2О) и углекислого газа (СО2) с использованием энергии солнечного света. Хлоропласты содержатся преимущественно в клетках листьев (у высших растений). Они сформированы двумя параллельно расположенными друг другу мембранами, окружающими содержимое хлоропластов — строму. Внутренняя мембрана образует многочисленные уплощенные мешочки — тилакоиды, которые сложены в стопки (наподобие стопки монет) — граны — и лежат в строме. Именно в тила-коидах и содержится хлорофилл.
Хромопласты определяют желтый, оранжевый и красный цвет многих цветков и плодов, в клетках которых присутствуют в большом количестве. Основными пигментами в их составе являются каротины. Функциональное назначение хромопластов состоит в цветовом привлечении животных, обеспечивающих опыление цветков и распространение семян.
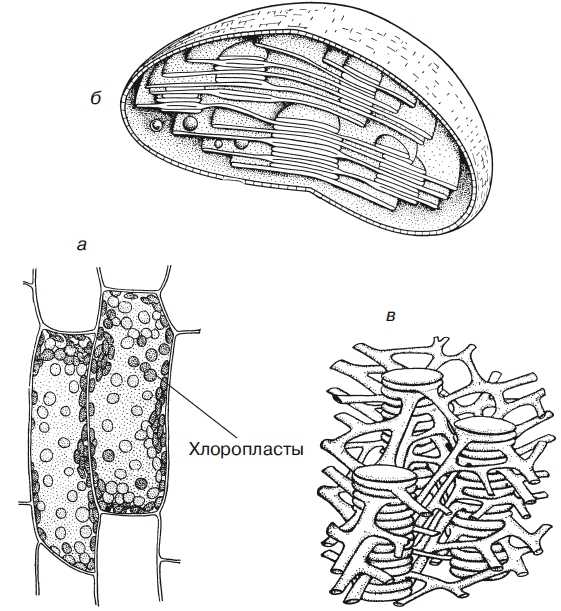
Рис. 2. Пластиды: а — хлоропласты в клетках листа элодеи, видимые в световом микроскопе; б — схема внутреннего строения хлоропласта с гранами, представляющими собой стопки плоских мешочков, расположенных перпендикулярно поверхности хлоропласта; в — более подробная схема, на которой видны анастомозирующие трубочки, соединяющие отдельные камеры гран
Лейкопласты - это бесцветные пластиды, содержащиеся в клетках подземных частей растений (например, в клубнях картофеля), семян и сердцевины стеблей. В лейкопластах, главным образом, происходит образование из глюкозы крахмала и накапливание его в запасающих органах растений.
Пластиды одного вида могут превращаться в другой. Например, при осеннем изменении цвета листьев хлоропласты превращаются в хромопласты.
Источник: Краснодембский Е. Г."Общая биология: Пособие для старшеклассников и поступающих в вузы"
xn--90aeobapscbe.xn--p1ai
 |
| Рисунок 1. Митохондрия под микроскопом. |
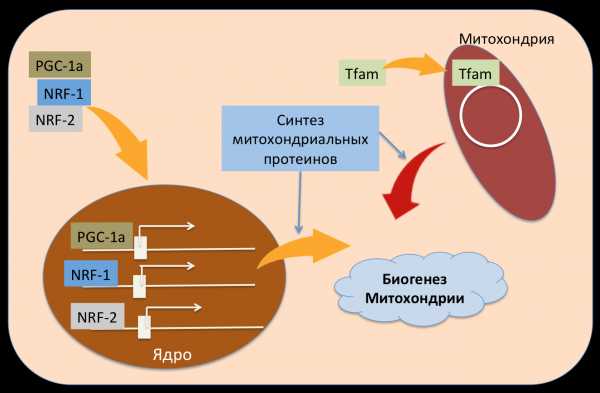
Но сегодня я буду рассказывать о другом немаловажном процессе - биогенез митохондрий (формирование новых митохондрий).Итак, биогенез митохондрии также очень сложный процесс и к сожалению не до конца изучен. Сложность заключается в том что в этом процессе участвуют более 1000 генов, около 20% всех клеточных протеинов изменяют свой уровень экспрессии во время этого процесса, и также биогенез митохондрии нуждается в слаженной кооперации двух геномов (клеточного и митохондриального).
Весь митохондриальный ДНК млекопитающих кодирует 13 протеинов участвующих в работе митохондрии (составные части дыхательной цепи переноса электронов), остальные протеины из которых состоит митохондрия кодируется ядерным ДНК а это ни много ни мало около 95% всех митохондриальных протеинов. Плюс ко всему транскрипция (считывание РНК) и репликация (удвоение ДНК) мхДНК происходит под жёстким контролем протеинов (кодирующиеся ядерным ДНК).Несмотря на всю сложность этого процесса, на сегодняшний день выделяют 3 протеина которые считаются "золотым стандартом" биогенеза митохондрий это Peroxisome proliferator-activated receptor gamma coactivator 1-alpha и -beta (PGC-1alpha и beta), Nuclear respiratory factors (NRF-1 и 2) и mitochondrial transcription factor A (Тfam).
 |
| Рисунок 2 |
Что нам скажут нокаут мыши?Мыши у которых отсутвтвует ген кодирующий NRF-1 или -2 нам ничего не скажут, так как они умирают во время эмбрионального развития (что подтверждает значимость биогенеза митохондрии в правильном функционировании организма). Такая же ситуация обстоит и с мышами у которых отсутвует Tfam ген (эмбрионально летальны). Когда как, PGC-1 alpha нокаут мыши в целом мало чем отличаются от обычных мышей, и даже митохондриальный биогенез на должном уровне, хотя многие исследования показывали важность этого протеина в биогенезе. Далее выяснилось что PGC-1 alpha дефицитные мыши экспрессируют в несколько раз больше PGC-1 beta, тем самым компенсируя недостаток своего alpha "собрата".
Для чего это нужно? Митохондрии "как и всё на свете" стареют что делает невозможным их функционирование, старые митохондрии утилизируются (благодаря таким процессам как аутофагия) и вместо них клетка формирует новые митохондрии. Также митохондрии очень чувствительны к различным клеточным стрессам (оксидативный стресс, воспалительный стимул, микробная инфекция итд) что приводит к повреждению митохондрии, и чтобы поддерживать энергетический баланс клетка запускает механизмы биогенеза новых митохондрии (Рисунок 3).
 |
| Рисунок3 |
Литература:Physiol Rev 88: 611–638, 2008Exp Gerontol. 2008 43(9): 813–819Mol Cell 14: 1–15, 2004Mol Cell Biol 21: 644–654, 2001Nature Genet 18: 231–236, 1998Am J Resp Crit Care Med. 2010;182:745-751.Am J Resp Crit Care Med. 2007;176:768-777.
naukageek.blogspot.com
МИТОХОНДРИИ (mitochondria; греч, mitos нить + chondrion зернышко) — органоиды, присутствующие в цитоплазме клеток животных и растительных организмов. М. принимают участие в процессах дыхания и окислительного фосфорилирования, продуцируют энергию, необходимую для функционирования клетки, представляя, таким образом, ее «силовые станции».
Термин «митохондрии» был предложен в 1894 г. Бендой (С. Benda). В середине 30-х гг. 20 в. удалось впервые выделить М. из клеток печени, что позволило исследовать эти структуры биохим, методами. В 1948 г. Хогебумом (G. Hogeboom) были получены окончательные доказательства того, что М. действительно являются центрами клеточного дыхания. Значительные успехи в изучении этих органоидов были сделаны в 60—70 гг. в связи с применением методов электронной микроскопии и молекулярной биологии.
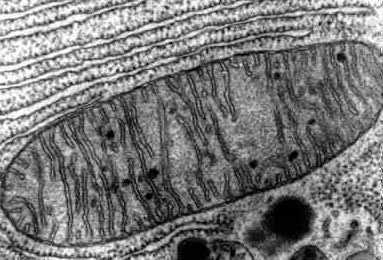
 Рис. 1. Электронограммы различных форм митохондрий из клетки поджелудочной железы крысы: а — удлиненная митохондрия; б — округлая митохондрия; X 38 000.
Рис. 1. Электронограммы различных форм митохондрий из клетки поджелудочной железы крысы: а — удлиненная митохондрия; б — округлая митохондрия; X 38 000. Форма М. варьирует от почти круглых до сильно вытянутых, имеющих вид нитей (рис. 1), Размер их колеблется от 0,1 до 7 мкм. Количество М. в клетке зависит от типа ткани и функционального состояния организма. Так, в сперматозоидах число М. невелико — ок. 20 (на клетку), в клетках эпителия почечных канальцев млекопитающих их содержится до 300 в каждой, а у гигантской амебы (Chaos chaos) обнаружено 500 000 митохондрий, В одной клетке печени крысы ок. 3000 М., однако в процессе голодания животного число М. может сократиться до 700. Обычно М. распределяются в цитоплазме достаточно равномерно, однако в клетках нек-рых тканей М. могут быть постоянно локализованы в участках, особенно нуждающихся в энергии. Напр., в скелетной мышце М. часто находятся в контакте с контрактильными участками миофибрилл, образуя правильные трехмерные структуры. В сперматозоидах М. образуют спиральный футляр вокруг осевой нити хвоста, что, вероятно, связано с возможностью использовать энергию АТФ, синтезируемую в М., для движений хвоста. В аксонах М. концентрируются вблизи синаптических окончаний, где происходит процесс передачи нервных импульсов, сопровождающийся энергозатратой. В клетках эпителия почечных канальцев М. связаны с выпячиваниями базальной клеточной мембраны. Это вызвано необходимостью постоянного и интенсивного снабжения энергией процесса активного переноса воды и растворенных в ней веществ, протекающего в почках.
 Рис. 2. Схема ультраструктуры митохондрии: а —трехмерное изображение митохондрии; б — тонкое строение митохондриальной кристы (соответствует участку, заключенному в рамку на рисунке a): 1 — наружная мембрана; 2 — внутренняя мембрана; 3 — матрикс; 4 — гранулы, содержащие ионы кальция и магния; 5 — кристы; 6 — F1-частицы; 7 — пространство между мембранами; молекулы белка (8) и липидов (9), образующие мембрану; 10 —цепи дыхательных ферментов, расположенные на наружной стороне внутренней мембраны.
Рис. 2. Схема ультраструктуры митохондрии: а —трехмерное изображение митохондрии; б — тонкое строение митохондриальной кристы (соответствует участку, заключенному в рамку на рисунке a): 1 — наружная мембрана; 2 — внутренняя мембрана; 3 — матрикс; 4 — гранулы, содержащие ионы кальция и магния; 5 — кристы; 6 — F1-частицы; 7 — пространство между мембранами; молекулы белка (8) и липидов (9), образующие мембрану; 10 —цепи дыхательных ферментов, расположенные на наружной стороне внутренней мембраны. Из особенностей строения внутренней мембраны необходимо отметить наличие в ней сферических частиц ок. 8—10 нм в поперечнике, сидящих на короткой ножке и иногда выступающих в матрикс. Эти частицы были открыты в 1962 г. Фернандес-Мораном (H. Fernandez-Moran). Они состоят из белка, обладающего АТФ-азной активностью, получившего обозначение F1. Белок прикрепляется к внутренней мембране только со стороны, обращенной к матриксу. Частицы F1 располагаются на расстоянии 10 нм друг от друга, а в каждой М. содержится 104—105, таких частиц.
В кристах и внутренних мембранах М. содержится большинство дыхательных ферментов (см.), дыхательные ферменты организованы в компактные ансамбли, распределенные с правильными промежутками в кристах М. на расстоянии 20 нм друг от друга.
М. почти всех типов клеток животных и растений построены по единому принципу, однако возможны отклонения в деталях. Так, кристы могут располагаться не только поперек длинной оси органоида, но и продольно, напр, в М. синаптической зоны аксона. В ряде случаев кристы могут ветвиться. В М. простейших организмов, нек-рых насекомых и в клетках клубочковой зоны надпочечников кристы имеют форму трубочек. Число крист различно; так, в М. клеток печени и половых клеток крист очень мало и они короткие, в то время как матрикс обилен; в М. мышечных клеток кристы многочисленны, а матрикса мало. Существует мнение, что число крист коррелирует с окислительной активностью М.
Во внутренней мембране М. осуществляются параллельно три процесса: окисление субстрата цикла Кребса (см. Трикарбоновых кислот цикл), перенос освободившихся при этом электронов и накопление энергии путем образования макроэргических связей аденозинтрифосфата (см. Аденозинфосфорные кислоты). Основной функцией М. является сопряжение синтеза АТФ (из АДФ и неорганического фосфора) и аэробного процесса окисления (см. Окисление биологическое). Накопленная в молекулах АТФ энергия может трансформироваться в механическую (в мышцах), электрическую (нервная система), осмотическую (почки) и т. д. Процессы аэробного дыхания (см. Окисление биологическое) и сопряженного с ним окислительного фосфорилирования (см.) являются основными функциями М. Кроме того, в наружной мембране М. может происходить окисление жирных к-т, фосфолипидов и нек-рых других соединений.
В 1963 г. Насс и Насс (М. Nass, S. Nass) установили, что в М. содержится ДНК (одна или несколько молекул). Все исследованные до сих пор митохондриальные ДНК из животных клеток состоят из ковалентно замкнутых колец диам. ок. 5 нм. У растений митохондриальная ДНК значительно длиннее и не всегда имеет форму кольца. Митохондриальная ДНК во многих отношениях отличается от ядерной. Репликация ДНК происходит при помощи обычного механизма, однако не совпадает во времени с репликацией ядерной ДНК. Количество генетической информации, заключенной в молекуле митохондриальной ДНК, по-видимому, недостаточно для кодирования всех белков и ферментов, содержащихся в М. Митохондриальные гены кодируют в основном структурные белки мембран и белки, участвующие в морфогенезе митохондрий. М. имеют свои транспортные РНК и синтетазы, содержат все компоненты, необходимые для синтеза белка; их рибосомы меньше цитоплазматических и более похожи на рибосомы бактерий.
Продолжительность жизни М. сравнительно невелика. Так, время обновления половины количества М. составляет для печени 9,6—10,2 сут., для почки — 12,4 сут. Пополнение популяции М. происходит, как правило, из предсуществующих (материнских) М. путем их деления или почкования.
Давно высказывалось предположение, что в процессе эволюции М. возникли, вероятно, путем эндосимбиоза примитивных ядросодержащих клеток с бактериоподобными организмами. Имеется большое число доказательств этому: наличие собственной ДНК, более сходной с ДНК бактерий, чем с ДНК ядра клетки; присутствие в М. рибосом; синтез ДНК-зависимой РНК; чувствительность митохондриальных белков к антибактериальному препарату — хлорамфениколу; сходство с бактериями в реализации дыхательной цепи; морфол., биохим, и физиол, различия между внутренней и наружной мембраной. Согласно симбиотической теории клетка-хозяин рассматривается как анаэробный организм, источником энергии для к-рого является гликолиз (протекающий в цитоплазме). В «симбионте» же реализуется цикл Кребса и дыхательная цепь; он способен к дыханию и окислительному фосфорилированию (см.).
М. являются весьма лабильными внутриклеточными органоидами, раньше других реагирующими на возникновение каких-либо патол, состояний. Возможны изменения числа М. в клетке (вернее, в их популяциях) или изменения их структуры. Напр., при голодании, действии ионизирующего облучения число М. уменьшается. Структурные изменения обычно состоят в набухании всего органоида, просветлении матрикса, разрушении крист, нарушении целостности наружной мембраны.
Набухание сопровождается значительным изменением объема М. В частности, при ишемии миокарда объем М. увеличивается в 10 раз и более. Различают два типа набухания: в одном случае оно связано с изменением осмотического давления внутри клетки, в других случаях — с изменениями клеточного дыхания, сопряженного с ферментативными реакциями и первичными функциональными расстройствами, вызывающими изменения водного обмена. Помимо набухания, может происходить вакуолизация М.
Независимо от причин, вызывающих патол, состояние (гипоксия, гиперфункция, интоксикация), изменения М. довольно стереотипны и неспецифичны.
Наблюдаются такие изменения структуры и функции М., к-рые, по-видимому, становились причиной возникновения болезни. В 1962 г. Луфт (R. Luft) описал случай «митохондриальной болезни». Больному с резко повышенной интенсивностью обмена веществ (при нормальной функции щитовидной железы) была сделана пункция скелетной мышцы и найдено повышенное число М., а также нарушение структуры крист. Дефектные митохондрии в клетках печени наблюдались и при выраженном тиреотоксикозе. Виноград (J. Vinograd) с сотр. (с 1937 по 1969) обнаружил, что у больных с определенными формами лейкемии митохондриальные ДНК из лейкоцитов заметно отличались от нормальных. Они представляли собой открытые кольца или группы сцепленных колец. Частота этих аномальных форм снижалась в результате химиотерапии.
См. также Клетка.
Библиография: Гаузе Г. Г. Митохондриальная ДНК, М., 1977,библиогр.; Д e P о-бертис Э., Новинский В. и С а э с Ф. Биология клетки, пер. с англ., М., 1973; Озернюк Н. Д. Рост и воспроизведение митохондрий, М., 1978, библиогр.; Поликар А. и Бесси М. Элементы патологии клетки, пер. с франц., М., 1970; РудинД. и Уилки Д. Биогенез митохондрий, пер. с англ., М., 1970, библиогр.; Серов В. В. и Пауков В. С. Ультраструктурная патология, М., 1975; С э д ж e р Р. Цитоплазматические гены и органеллы, пер. с англ., М., 1975.
Т. А. Залетаева.
xn--90aw5c.xn--c1avg
| Митохондрии — «силовые» станции клетки, в них локализована большая часть реакций дыхания (аэробная фаза). В митохондриях происходит аккумуляция энергии дыхания в аденозинтрифосфате (АТФ). Энергия, запасаемая в АТФ, служит основным источником для физиологической деятельности клетки. Митохондрии обычно имеют удлиненную палочковидную форму длиной 4—7 мкм и диаметром 0,5—2 мкм. Число митохондрий в клетке может быть различным от 500 до 1000 и зависит от роли данного органа в процессах энергетического обмена. Однако в некоторых организмах (дрожжах) имеется лишь одна гигантская митохондрия.
Электронно- микроскопическая фотография митохондрий (по П. В. Парамоновой) Химический состав митохондрий несколько колеблется. В основном это белково-липидные органеллы. Содержание белка в них составляет 60—65%, причем структурные и ферментативные белки содержатся примерно в равной пропорции, а также около 30% липидов. Очень важно, что митохондрии содержат нуклеиновые кислоты: РНК — 1 % и ДНК —0,5%. В митохондриях имеется не только ДНК, но и вся система синтеза белка, в том числе рибосомы. Митохондрии окружены двойной мембраной. Толщина мембран составляет 6—10 нм. Мембраны митохондрий на 70% состоят из белка. Фосфолипиды мембран представлены фосфатидтилхолином, фосфатидилэтаноламином, а также специфическими фосфолипидами, например, кардиолипином. Мембраны митохондрий не пропускают Н+ и служат барьером для их транспорта. Это важнейшее свойство лежит в основе образования электрохимического градиента. Между мембранами находится заполненное жидкостью перимитохондриальное пространство, равное 10 нм. Внутреннее пространство митохондрий заполняет матрикс в виде студнеобразной полужидкой массы. В матриксе сосредоточены ферменты цикла Кребса. Внутренняя мембрана дает выросты — кристы, расположенные перпендикулярно продольной оси органеллы и перегораживающие все внутреннее пространство митохондрий на отдельные отсеки. Однако, поскольку выросты-перегородки неполные, между этими отсеками сохраняется связь. Мембраны митохондрий обладают большой прочностью и гибкостью. Во внутренней мембране локализована дыхательная цепь (цепь переноса электронов). На внутренней поверхности внутренней мембраны митохондрий равномерно расположены грибовидные частицы. Каждая митохондрия содержит 104—105 таких грибовидных частиц, которые представляют собой фермент АТФ-синтазу, катализирующую образование АТФ.Митохондрии способны к движению. Это имеет большое значение в жизни клетки, так как митохондрии передвигаются к тем местам, где идет усиленное потребление энергии. Они могут ассоциировать друг с другом, как путем тесного сближения, так и при помощи связующих тяжей. Наблюдаются также контакты митохондрий с эндоплазматической сетью, ядром, хлоропластами. Известно, что митохондрии способны к набуханию, а при потере воды — к уменьшению объема. В растущих клетках в митохондриальном матриксе количество крист растет — это коррелирует с увеличением интенсивности дыхания. В процессе дыхания ультраструктура митохондрий меняется. В том случае, если в митохондриях протекает активный процесс преобразования энергии окисления в энергию АТФ, внутренняя часть митохондрий становится более компактной. Митохондрии имеют свой онтогенез. В меристематических клетках можно наблюдать инициальные частицы, которые представляют собой округлые образования, окруженные двойной мембраной. Диаметр таких инициальных частиц составляет 50 нм. По мере роста клетки инициальные частицы увеличиваются в размере, удлиняются и их внутренняя мембрана образует выросты, перпендикулярные оси митохондрий. Вначале образуются промитохондрии. Они еще не достигают окончательного размера и имеют мало крист. Из промитохондрии образуются митохондрии. Сформировавшиеся митохондрии делятся путем перетяжки или почкованием.
Схема строения митохондрии: 1 — наружная мембрана; 3 — матрикс; 2 — внутренняя мембрана; 4 — перимитохондриальное пространство. Свойства митохондрий (белки, структура) закодированы частично в ДНК митохондрий, а частично в ядре. Так, митохондриальный геном кодирует белки рибосом и частично систему переносчиков электронотранспортной цепи, а в геноме ядра кодирована информация о белках-ферментах цикла Кребса. Сопоставление размеров митохондриальной ДНК с числом и размером мито-хондриальных белков показывает, что в ней заложено информации почти для половины белков. Это и позволяет считать митохондрии, как и хлоропласты, полуавтономными, т. е. не полностью зависящими от ядра. Они имеют собственную ДНК и собственную белоксинтезирующую систему, и именно с ними и с пластидами связана так называемая цитоплазматическая наследственность. В большинстве случаев это наследование по материнской линии, так как инициальные частицы митохондрий локализованы в яйцеклетке. Таким образом, митохондрии всегда образуются от митохондрий. Широко обсуждается вопрос, как рассматривать митохондрии и хлоропласты с эволюционной точки зрения. Еще в 1921 г. русский ботаник Б.М. Козо-Полянский высказал мнение, что клетка — это симбиотрофная система, в которой сожительствует несколько организмов. В настоящее время эндосимбиотическая теория происхождения митохондрий и хлоропластов является общепринятой. Согласно этой теории, митохондрии — это в прошлом самостоятельные организмы. По мнению Л. Маргелис (1983), это могли быть эубактерии, содержащие ряд дыхательных ферментов. На определенном этапе эволюции они внедрились в примитивную, содержащую ядро, клетку. Оказалось, что ДНК митохондрий и хлоропластов по своей структуре резко отличается от ядерной ДНК высших растений и сходна с бактериальной ДНК (кольцевое строение, нуклеотидная последовательность). Сходство обнаруживается и по величине рибосом. Они мельче цитоплазматических рибосом. Синтез белка в митохондриях, подобно бактериальному, подавляется антибиотиком хлорамфениколом, который не влияет на синтез белка на рибосомах эукариот. Кроме того, система переноса электронов у бактерий расположена в плазматической мембране, что напоминает организацию электронтранспортной цепи во внутренней митохондриальной мембране. |
fizrast.ru
Цитоплазма. Отграниченная от внешней среды плазматической мембраной, цитоплазма представляет собой внутреннюю полужидкую среду клеток. В цитоплазме эукариотических клеток располагаются ядро и различные органоиды. Ядро располагается в центральной части цитоплазмы. В ней сосредоточены и разнообразные включения – продукты клеточной деятельности, вакуоли, а также мельчайшие трубочки и нити, образующие скелет клетки. В составе основного вещества цитоплазмы преобладают белки. В цитоплазме протекают основные процессы обмена веществ, она объединяет в одно целое ядро и все органоиды, обеспечивает их взаимодействие, деятельность клетки как единой целостной живой системы.
Эндоплазматическая сеть. Вся внутренняя зона цитоплазмы заполнена многочисленными мелкими каналами и полостями, стенки которых представляют собой мембраны, сходные по своей структуре с плазматической мембраной. Эти каналы ветвятся, соединяются друг с другом и образуют сеть, получившую название эндоплазматической сети [55], [56]. Рис. 55. Электронно-микроскопическая фотография ядра и участка эндоплазматической сети, расположенной в цитоплазме (Видны каналы и более крупные полости эндоплазматической сети, митохондрии овальной формы. Видны также ядерная оболочка с порами, ядерный сок ядрышка. Увел. 20 000.)
Рис. 55. Электронно-микроскопическая фотография ядра и участка эндоплазматической сети, расположенной в цитоплазме (Видны каналы и более крупные полости эндоплазматической сети, митохондрии овальной формы. Видны также ядерная оболочка с порами, ядерный сок ядрышка. Увел. 20 000.) Рис. 56. Электронно-микроскопическая фотография гранулярной эндоплазматической сети с рибосомами (округлые темные тельца) на поверхности ее мембран. Увел. 70 000.Эндоплазматическая сеть неоднородна по своему строению. Известны два ее типа – гранулярная [56] и гладкая [57]. На мембранах каналов и полостей гранулярной сети располагается множество мелких округлых телец рибосом [56], которые придают мембранам шероховатый вид. Мембраны гладкой эндоплазматической сети не несут рибосом на своей поверхности.
Рис. 56. Электронно-микроскопическая фотография гранулярной эндоплазматической сети с рибосомами (округлые темные тельца) на поверхности ее мембран. Увел. 70 000.Эндоплазматическая сеть неоднородна по своему строению. Известны два ее типа – гранулярная [56] и гладкая [57]. На мембранах каналов и полостей гранулярной сети располагается множество мелких округлых телец рибосом [56], которые придают мембранам шероховатый вид. Мембраны гладкой эндоплазматической сети не несут рибосом на своей поверхности. Рис. 57. Гладкая эндоплазматическая сетьЭндоплазматическая сеть выполняет много разнообразных функций. Основная функция гранулярной эндоплазматической сети – участие в синтезе белка, который осуществляется в рибосомах.
Рис. 57. Гладкая эндоплазматическая сетьЭндоплазматическая сеть выполняет много разнообразных функций. Основная функция гранулярной эндоплазматической сети – участие в синтезе белка, который осуществляется в рибосомах.
На мембранах гладкой эндоплазматической сети происходит синтез липидов и углеводов. Все эти продукты синтеза накапливаются в каналах и полостях, а затем транспортируются к различным органоидам клетки, где потребляются или накапливаются в цитоплазме в качестве клеточных включений. Эндоплазматическая сеть связывает между собой основные органоиды клетки [55].
Рибосомы. Рибосомы обнаружены в клетках всех организмов. Это микроскопические тельца округлой формы диаметром 15– 20 нм. Каждая рибосома состоит из двух неодинаковых по размерам частиц, малой и большой, как это показано на рисунке 58.Рис. 58. Электронно-микроскопическая фотография полисомы и схема полисомы и рибосомыВ одной клетке содержится много тысяч рибосом, они располагаются либо на мембранах гранулярной эндоплазматической сети, либо свободно лежат в цитоплазме [56].
В состав рибосом входят белки и РНК.
Функция рибосом – это синтез белка. Синтез белка сложный процесс, который осуществляется не одной рибосомой, а целой группой, включающей до нескольких десятков объединенных рибосом. Такую группу рибосом называют полисомой [58].
Синтезированные белки сначала накапливаются в каналах и полостях эндоплазматической сети, а затем транспортируются к органоидам и участкам клетки, где они потребляются. Эндоплазматическая сеть н рибосомы, расположенные на ее мембранах, представляют собой единый аппарат биосинтеза и транспортировки белков.
Митохондрии. В цитоплазме большинства клеток животных и растений содержатся мелкие тельца (0,2 – 7 мкм) – митохондрии (греч. «митос» – нить, «хондрнон» – зерно, гранула; [50], [59]). Рис. 59. Электронно-микроскопическая фотография митохондрии. Видны наружная и внутренняя мембрана и кристы. Увел. 40 000.Митохондрии хорошо видны в световой микроскоп, с помощью которого можно рассмотреть их форму, расположение, сосчитать количество. Внутреннее стояние митохондрий изучено с помощью электронного микроскопа [60] . Как видно на рисунке, оболочка митохондрии состоит из двух мембран – наружной и внутренней. Наружная мембрана гладкая, она не образует никаких складок и выростов. Внутренняя мембрана, напротив, образует многочисленные складки, которые направлены в полость митохондрии [60]. Складки внутренней мембраны называют кристами (лат. «криста» – гребень, вырост). Число крист неодинаково в митохондриях разных клеток. Их может быть от нескольких десятков до нескольких сотен, причем особенно много крист в митохондриях активно функционирующих клеток, например мышечных.Рис. 60. Схема строения митохондрии по данным электронного микроскопаМитохондрии называют «силовыми станциями» клеток, так как их основная функция -. синтез аденозинтрифосфорной кислоты (АТФ). Эта кислота синтезируется в митохондриях клеток всех организмов и представляет собой универсальный источник энергии, необходимый для осуществления процессов жизнедеятельности клетки и целого организма.
Рис. 59. Электронно-микроскопическая фотография митохондрии. Видны наружная и внутренняя мембрана и кристы. Увел. 40 000.Митохондрии хорошо видны в световой микроскоп, с помощью которого можно рассмотреть их форму, расположение, сосчитать количество. Внутреннее стояние митохондрий изучено с помощью электронного микроскопа [60] . Как видно на рисунке, оболочка митохондрии состоит из двух мембран – наружной и внутренней. Наружная мембрана гладкая, она не образует никаких складок и выростов. Внутренняя мембрана, напротив, образует многочисленные складки, которые направлены в полость митохондрии [60]. Складки внутренней мембраны называют кристами (лат. «криста» – гребень, вырост). Число крист неодинаково в митохондриях разных клеток. Их может быть от нескольких десятков до нескольких сотен, причем особенно много крист в митохондриях активно функционирующих клеток, например мышечных.Рис. 60. Схема строения митохондрии по данным электронного микроскопаМитохондрии называют «силовыми станциями» клеток, так как их основная функция -. синтез аденозинтрифосфорной кислоты (АТФ). Эта кислота синтезируется в митохондриях клеток всех организмов и представляет собой универсальный источник энергии, необходимый для осуществления процессов жизнедеятельности клетки и целого организма.
Новые митохондрии образуются делением уже существующих в клетке митохондрий.
Пластиды. В цитоплазме клеток всех растений находятся пластиды. В клетках животных пластиды отсутствуют. Различают три основных типа пластид: зеленые в хлоропласты; красные, оранжевые и желтые в хромопласты; бесцветные лейкопласты.
Хлоропласты. Эти органоиды содержатся в клетках листьев и других зеленых органов растений, а также у разнообразных водорослей. Размеры хлоропластов 4 – 6 мкм, наиболее часто они имеют овальную форму. У высших растений в одной клетке обычно бывает несколько десятков хлоропластов. Зеленый цвет хлоропластов зависит от содержания в них пигмента хлорофилла. Хлоропласт – основной органоид клеток растений, в котором происходит фотосинтез, т. е. образование органических веществ (углеводов) из неорганических (CO2 и h3O) при использовании энергии солнечного света.
По строению хлоропласты сходны с митохондриями. От цитоплазмы хлоропласт отграничен двумя мембранами – наружной и внутренней [61]. Наружная мембрана гладкая, без складок и выростов, а внутренняя образует много складчатых выростов, направленных внутрь хлоропласта. Поэтому внутри хлоропласта сосредоточено большое количество мембран, образующих особые структуры – граны. Они сложены наподобие стопки монет [61]. Рис. 61. Электронно-микроскопическая фотография хлоропласта. Увел. 40 000.В мембранах гран располагаются молекулы хлорофилла, потому именно здесь происходит фотосинтез. В хлоропластах синтезируется и АТФ. Между внутренними мембранами хлоропласта содержатся ДНК, РНК и рибосомы. Следовательно, в хлоропластах, так же как и в митохондриях, происходит синтез белка, необходимого для деятельности этих органоидов. Хлоропласты размножаются делением.
Рис. 61. Электронно-микроскопическая фотография хлоропласта. Увел. 40 000.В мембранах гран располагаются молекулы хлорофилла, потому именно здесь происходит фотосинтез. В хлоропластах синтезируется и АТФ. Между внутренними мембранами хлоропласта содержатся ДНК, РНК и рибосомы. Следовательно, в хлоропластах, так же как и в митохондриях, происходит синтез белка, необходимого для деятельности этих органоидов. Хлоропласты размножаются делением.
Хромопласты находятся в цитоплазме клеток разных частей растений: в цветках, плодах, стеблях, листьях. Присутствием хромопластов объясняется желтая, оранжевая и красная окраска венчиков цветков, плодов, осенних листьев.
Лейкопласты бесцветны. Они содержатся в цитоплазме неокрашенных частей растений, например в стеблях, корнях, клубнях. Форма лейкопластов разнообразна. Примером широко распространенных лейкопластов могут служить лейкопласты клубней картофеля, в которых накапливаются зерна крахмала.
Хлоропласты, хромопласты и лейкопласты способны к взаимному переходу. Так, при созревании плодов или изменении окраски листьев осенью хлоропласты превращаются в хромопласты, а лейкопласты могут превращаться в хлоропласты, например, при позеленении клубней картофеля.
1. На рисунке 50 найдите органоиды, расположенные в цитоплазме. 2. Каковы строение и функции эндоплазматической сети, рибосом, митохондрий, хлоропластов? 3. Какие типы пластид находятся в растительной клетке и какова их роль? 4. Сравните функции митохондрий и хлоропластов.
blgy.ru
Существует гипотеза о том, что в известной степени автономные и несущие определенное количество собственной ДНК митохондрии и пластиды представляют собой видоизмененные прокариотические организмы, которые нашли "убежище" в более крупных гетеротрофных клетках-хозяевах - предшественниках эукариот . Все, или почти все, ныне живущие эукариоты содержат в своих клетках митохондрии, а все автотрофные эукариоты - также и пластиды . Возможно, они были приобретены в результате независимых случаев симбиоза , точнее - внутреннего симбиоза (эндосимбиоза). Более крупные гетеротрофные клетки, предшественницы эукариотических клеток , очевидно, защищали свои симбиотические органоиды от различных неблагоприятных условий окружающей среды. В свою очередь, прокариотические симбионты оказались полезными благодаря способности использовать энергию солнечного света ( фотосинтез ) и возможности использовать молекулярный кислород для окисления органических веществ. В результате эукариоты смогли заселить сушу, а также ту часть водной среды (обычно с высокой рН), где прокариоты относительно немногочисленны.
Митохондрии - неотъемлемая часть всех живых эукариотических клеток. Форма, величина и их число постоянно меняются. Число митохондрий варьирует от нескольких десятков до сотен. Особенно их много в секреторных тканях растений. Размеры этих органоидов не превышают 1 мкм. По форме они чаще всего эллиптические или округлые. Снаружи митохондрии окружены оболочкой, состоящей из двух мембран, которые не связаны с эндоплазматической сетью цитоплазмы ( рис. 9 ). Внутренняя мембрана образует выросты в полость митохондрии в виде пластин или трубок, называемых кристами . Кристы бывают различных типов. Пространство между кристами заполнено однородным прозрачным веществом - матриксом митохондрий. В матриксе встречаются рибосомы , подобные по величине рибосомам прокариотических клеток, и собственная митохондриальная ДНК , заметная под электронным микроскопом в виде тонких нитей.
Митохондрии способны к независимому от ядра синтезу своих белков на собственных рибосомах под контролем митохондриальной ДНК . Митохондрии образуются только путем деления.
Основная функция митохондрий состоит в обеспечении энергетических потребностей клетки путем дыхания. Богатые энергией молекулы АТФ синтезируются при реакции окислительного фосфорилирования. Энергия, запасаемая АТФ, получается в результате окисления в митохондриях различных энергетически богатых веществ, главным образом сахаров . Механизм окислительного фосфорилирования путем хемиосмотического сопряжения открыт в 1960 г. английским биохимиком П.Митчеллом.
medbiol.ru